
In this blog, we will present internal data and information on priming RoosterBio’s bone marrow-derived human MSCs (hMSC) with pro-inflammatory molecules and the impact of these priming protocols on hMSC immunomodulatory function and angiogenic cytokine secretion.
Introduction
Human Mesenchymal Stem/Stromal Cells, or hMSCs, are key components of future therapeutics, engineered tissues, and medical devices. There are currently over 400 clinical trials investigating hMSCs as therapies (1). The trials have produced some promising results, with hMSCs generally deemed safe, and in some cases effective (2). It is believed that these versatile cells achieve their biologic and therapeutic effects by secreting a plethora of biomolecules (referred to as the MSC secretome) that moderate a variety of processes including angiogenesis, immunosuppression, and overall “tissue repair” (3-6). As the secretome is one of the likely Mechanisms of Actions (MOA) of hMSC therapies, there is a significant amount of recent work on engineering hMSC preparations to enhance secreted factors by genetic modification, by culture strategies, or by engineering the hMSC microenvironment (5, 7-11). In addition, a recent ISCT paper (12) advances the concept of “priming” hMSCs by exposing the cells to pro-inflammatory cytokines prior to implantation.
Thus, priming of hMSCs can have two primary purposes:
- To assess human MSC preparations in vitro as recommended by the ISCT and the FDA (12, 13), and
- To enhance hMSC potency (survival, immunosuppression, homing) prior to implantation (8, 9, 14).
As part of our standard quality control (QC) testing, RoosterBio analyzes the immunosuppressive capability of our cell lots through priming with IFN-γ. The hMSC response (i.e. immunomodulatory potential) is reported as a measure of indoleamine 2,3-dioxygenase (IDO) activity, determined by measuring the amino acid kynurenine in the culture supernatant. The IDO enzyme converts L-tryptophan to N-formylkynurenine (or kynurenine), an immunosuppressive molecule that acts as an inhibitor of immune cell proliferation – including T cells (12, 15, 16). Testing every hMSC lot for inducible IDO activity provides a quality assurance that the cells we release have some level of functional potency as it relates to immunomodulation– which we consider a key quality attribute of hMSCs.
While researchers are beginning to implement testing of hMSC preparations for inducible IDO activity prior to implantation, few are looking at the impact of priming on other hMSC functions. Here, we present information on priming of RoosterBio’s hMSCs with IFN-γ ± TNF-α across multiple lots and donors and the impact of such treatment on hMSC IDO activity and angiogenic cytokine secretion. The goal of this blog post is to demonstrate that priming has impacts on several functional properties of hMSCs, and that researchers should consider priming regimens to (a) understand the potency of their specific cell products, especially in inflammatory environments, and (b) to potentially increase potency of these cell products upon therapeutic administration.
Methods & Experimental Design
Materials & Reagents
Cell culture reagents were purchased from Life Technologies, chemicals and reagents for kynurenine measurement were from Sigma, and cultureware was from Corning. Other products are: RoosterVial™ (BM-hMSC, part # MSC-001, RoosterBio) and RoosterBio RoosterNourish™-MSC (SKU KT-001).
hMSC Priming
RoosterBio bone marrow-derived hMSCs from multiple manufacturing cell lots and several donors were used for these experiments. All testing was performed on hMSCs of PDL range 10-14.
- Frozen hMSC vials were thawed and expanded in fully supplemented RoosterBio RoosterNourish-MSC according to the RoosterBio customer expansion protocol. Briefly, cells were plated in a T75 flask at 3000-3500 viable cells/cm2, incubated with no media change for 5-6 days, and harvested using TrypLE (LifeTech).
- For priming studies, hMSCs were seeded at confluence at 40,000 cells/cm2 in 12- or 6-well plates (Falcon) in 1 or 2 ml RoosterNourish. After 18-24 hr, media was changed, to exclude RoosterBio RoosterBooster™, by replacing media with RoosterBio basal media (SU-005) +2% FBS or DMEM +2% FBS.
- After 1 hr, cells were (a) left alone as a non-primed control, or were treated with (b) Interferon-γ (IFN-γ, LifeTech), at a final concentration of 10 ng/ml, (c) Tumor Necrosis Factor-α (TNF-α, LifeTech), at 10 ng/ml, or (d) IFN-γ and TNF-α together (10 ng/ml each).
- After 24 ± 1hr, culture supernatant was collected and frozen at -80˚C until assayed for IDO activity and cytokine secretion.
Measuring of Priming (IDO activity and Cytokines)
IDO activity measurement.
IDO activity was assayed by quantifying kynurenine in the culture supernatant using a published colorimetric assay (16-19).
- N-formylkynurenine was hydrolyzed to kynurenine by trichloroacetic acid (TCA) and treated with DMBA (1% (w/v) p-dimethylaminobenzaldehyde in acetic acid).
- Kynurenine was measured by absorbance at 480 nm. A standard curve of 0-10 μg/ml L- kynurenine (Sigma) in media was used to calculate kynurenine levels.
- IDO activity rates were calculated by normalizing kynurenine levels to the number of cells in the well and days of incubation. IDO activity is expressed as pg kynurenine/cell/day.
Cytokine secretion.
- Culture supernatant was collected and assayed for basic FGF (bFGF, FGF2), HGF, IL-8, TIMP-1, TIMP-2 and VEGF by MultiPlex ELISA (Quansys, Angiogenesis Panel).
- Secreted cytokine concentration was calculated and normalized to initial number of cells seeded and days of incubation to obtain specific cytokine secretion rates (presented as pg/cell/day).
Results
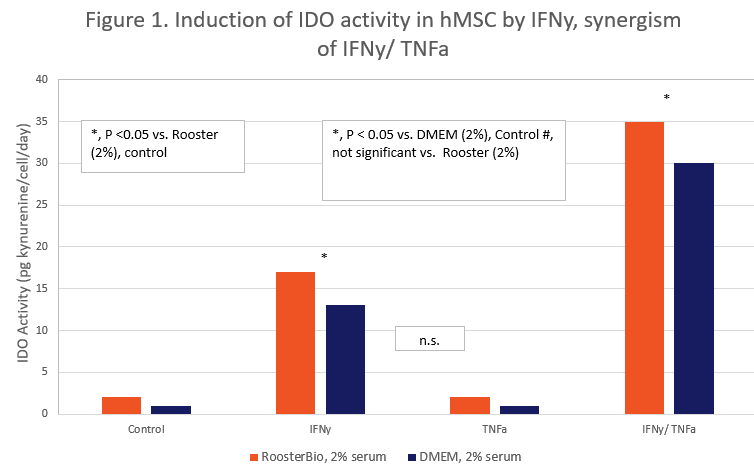
Priming can be tested in different basal media. Data presented in Figure 1 show that (a) hMSC can be primed in RoosterBio basal medium or DMEM and (b) that the up-regulation of IDO activity between cell lots and different hMSC donors is highly consistent, demonstrated by the small SEM error bars.
IFN-γ and TNF-α demonstrate synergism in hMSC priming. Data in Figure 1 also show that the IDO response of hMSC to IFN-γ + TNF-α is synergistic, which is consistent with other publications with hMSCs; i.e. hMSCs responding to IFN-γalone, no response to TNF-α alone, and a synergistic response to IFN-γ and TNF-α together (12, 20-22).
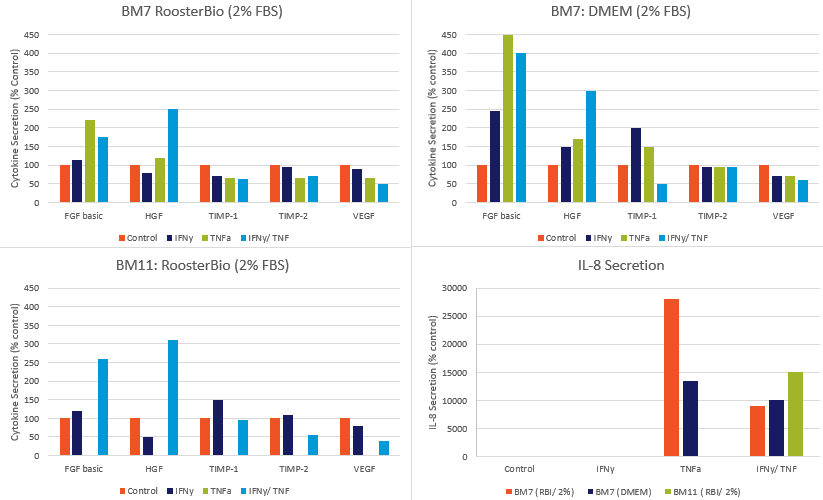
Angiogenic cytokine secretion is also variably regulated during priming. When angiogenic cytokine secretion was assayed under priming conditions, we found a complicated pattern of secretion (Figure 2). None of the angiogenic factors were greatly altered (>30% change up or down) by IFN-γ alone. Basic FGF (FGF2) increased with TNF-α; this change was IFN-γ independent. HGF increased with IFN-γ/TNF-α co-treatment, but not with either treatment alone. There was a very large increase in IL-8 secretion with TNF-α treatment. Interestingly IL-8 was decreased by IFN-γ (+/- TNF-α). There was a slight decrease in secretion of VEGF, an important trophic factor, with IFN-γ/TNF-α co-treatment. We have only performed cytokine testing across 2 donors, but consistent patterns were seen across lots in both RoosterBio basal medium and DMEM basal media.
Conclusions
Here, we present information on priming of RoosterBio hMSCs across various lots and donors. The current data demonstrates that:
- hMSCs from multiple donors can be primed in multiple basal media types.
- The effect of IDO induction by IFN-γ is synergistically amplified by co-treatment with TNF-α.
- There are complicated changes in hMSC angiogenic cytokine secretion with priming.
These results reinforce the need for more complete (global) expression studies of any treatments to hMSCs. In addition, biological results need to be interpreted carefully, and any biological effects of priming cannot be solely attributed to changes in immunomodulatory cytokines/chemokines as they may be a function of hMSC cytokine secretion as well.
The take-home message is that hMSC functional properties can be altered in vitro in response to various stimuli. Monitoring the various hMSC biological functions is an important part of quality control testing on final cell products. As demonstrated here, the simple addition of 2 factors can greatly alter hMSC function. Clearly, introducing cells into a living host will provide many more micro-environmental cues that will modulate hMSC response and function upon therapeutic administration. Extensive in vivo testing, and eventual clinical testing, will be required to understand if priming of hMSCs prior to delivery to the patient is a robust approach to enhance product potency. Thus, the impact of priming of hMSCs may go beyond simply using the approach as a standardization assay.
Readers, any comments or questions?
1. Li MD, Atkins H, & Bubela T (2014) The global landscape of stem cell clinical trials. Regenerative medicine 9(1):27-39. http://www.ncbi.nlm.nih.gov/pubmed/24236476
2. Lalu MM, McIntyre L, Pugliese C, Fergusson D, Winston BW, Marshall JC, Granton J, Stewart DJ, & Canadian Critical Care Trials G (2012) Safety of cell therapy with mesenchymal stromal cells (SafeCell): a systematic review and meta-analysis of clinical trials. PloS one 7(10):e47559. http://www.ncbi.nlm.nih.gov/pubmed/23133515
3. Murphy MB, Moncivais K, & Caplan AI (2013) Mesenchymal stem cells: environmentally responsive therapeutics for regenerative medicine. Experimental & molecular medicine 45:e54. http://www.ncbi.nlm.nih.gov/pubmed/24232253
4. Phinney DG, Galipeau J, Krampera M, Martin I, Shi Y, & Sensebe L (2013) MSCs: science and trials. Nature medicine 19(7):812. http://www.ncbi.nlm.nih.gov/pubmed/23836216
5. Ranganath SH, Levy O, Inamdar MS, & Karp JM (2012) Harnessing the mesenchymal stem cell secretome for the treatment of cardiovascular disease. Cell stem cell10(3):244-258. http://www.ncbi.nlm.nih.gov/pubmed/22385653
6. Baraniak PR & McDevitt TC (2010) Stem cell paracrine actions and tissue regeneration. Regenerative medicine 5(1):121-143. http://www.ncbi.nlm.nih.gov/pubmed/20017699
7. Betancourt AM (2013) New Cell-Based Therapy Paradigm: Induction of Bone Marrow-Derived Multipotent Mesenchymal Stromal Cells into Pro-Inflammatory MSC1 and Anti-inflammatory MSC2 Phenotypes. Advances in biochemical engineering/biotechnology 130:163-197. http://www.ncbi.nlm.nih.gov/pubmed/22869086
8. Luo Y, Wang Y, Poynter JA, Manukyan MC, Herrmann JL, Abarbanell AM, Weil BR, & Meldrum DR (2012) Pretreating mesenchymal stem cells with interleukin-1beta and transforming growth factor-beta synergistically increases vascular endothelial growth factor production and improves mesenchymal stem cell-mediated myocardial protection after acute ischemia. Surgery 151(3):353-363. http://www.ncbi.nlm.nih.gov/pubmed/22088815
9. Waterman RS, Henkle SL, & Betancourt AM (2012) Mesenchymal stem cell 1 (MSC1)-based therapy attenuates tumor growth whereas MSC2-treatment promotes tumor growth and metastasis. PloS one 7(9):e45590. http://www.ncbi.nlm.nih.gov/pubmed/23029122
10. Waterman RS, Tomchuck SL, Henkle SL, & Betancourt AM (2010) A new mesenchymal stem cell (MSC) paradigm: polarization into a pro-inflammatory MSC1 or an Immunosuppressive MSC2 phenotype. PloS one 5(4):e10088. http://www.ncbi.nlm.nih.gov/pubmed/20436665
11. Sart S, Tsai AC, Li Y, & Ma T (2013) Three-dimensional Aggregates of Mesenchymal Stem Cells: Cellular Mechanisms, Biological Properties, and Applications. Tissue engineering. Part B, Reviews. http://www.ncbi.nlm.nih.gov/pubmed/24168395
12. Krampera M, Galipeau J, Shi Y, Tarte K, & Sensebe L (2013) Immunological characterization of multipotent mesenchymal stromal cells–The International Society for Cellular Therapy (ISCT) working proposal. Cytotherapy15(9):1054-1061. http://www.ncbi.nlm.nih.gov/pubmed/23602578
13. Mendicino M, Bailey AM, Wonnacott K, Puri RK, & Bauer SR (2014) MSC-based product characterization for clinical trials: an FDA perspective. Cell stem cell14(2):141-145. http://www.ncbi.nlm.nih.gov/pubmed/24506881
14. Waterman RS, Morgenweck J, Nossaman BD, Scandurro AE, Scandurro SA, & Betancourt AM (2012) Anti-inflammatory mesenchymal stem cells (MSC2) attenuate symptoms of painful diabetic peripheral neuropathy. Stem cells translational medicine1(7):557-565. http://www.ncbi.nlm.nih.gov/pubmed/23197860
15. Francois M & Galipeau J (2012) New insights on translational development of mesenchymal stromal cells for suppressor therapy. Journal of cellular physiology227(11):3535-3538. http://www.ncbi.nlm.nih.gov/pubmed/22378308
16. Carmen J, Burger SR, McCaman M, & Rowley JA (2012) Developing assays to address identity, potency, purity and safety: cell characterization in cell therapy process development. Regenerative medicine 7(1):85-100. http://www.ncbi.nlm.nih.gov/pubmed/22168500
17. Daubener W, Wanagat N, Pilz K, Seghrouchni S, Fischer HG, & Hadding U (1994) A new, simple, bioassay for human IFN-gamma. Journal of immunological methods168(1):39-47. http://www.ncbi.nlm.nih.gov/pubmed/8288893
18. Littlejohn TK, Takikawa O, Truscott RJ, & Walker MJ (2003) Asp274 and his346 are essential for heme binding and catalytic function of human indoleamine 2,3-dioxygenase. The Journal of biological chemistry 278(32):29525-29531. http://www.ncbi.nlm.nih.gov/pubmed/12766158
19. Takikawa O, Kuroiwa T, Yamazaki F, & Kido R (1988) Mechanism of interferon-gamma action. Characterization of indoleamine 2,3-dioxygenase in cultured human cells induced by interferon-gamma and evaluation of the enzyme-mediated tryptophan degradation in its anticellular activity. The Journal of biological chemistry 263(4):2041-2048. http://www.ncbi.nlm.nih.gov/pubmed/3123485
20. Croitoru-Lamoury J, Lamoury FM, Caristo M, Suzuki K, Walker D, Takikawa O, Taylor R, & Brew BJ (2011) Interferon-gamma regulates the proliferation and differentiation of mesenchymal stem cells via activation of indoleamine 2,3 dioxygenase (IDO). PloS one 6(2):e14698. http://www.ncbi.nlm.nih.gov/pubmed/21359206
21. Krampera M (2011) Mesenchymal stromal cell ‘licensing’: a multistep process. Leukemia25(9):1408-1414. http://www.ncbi.nlm.nih.gov/pubmed/21617697
22. Meisel R, Brockers S, Heseler K, Degistirici O, Bulle H, Woite C, Stuhlsatz S, Schwippert W, Jager M, Sorg R, Henschler R, Seissler J, Dilloo D, & Daubener W (2011) Human but not murine multipotent mesenchymal stromal cells exhibit broad-spectrum antimicrobial effector function mediated by indoleamine 2,3-dioxygenase. Leukemia 25(4):648-654. http://www.ncbi.nlm.nih.gov/pubmed/21242993